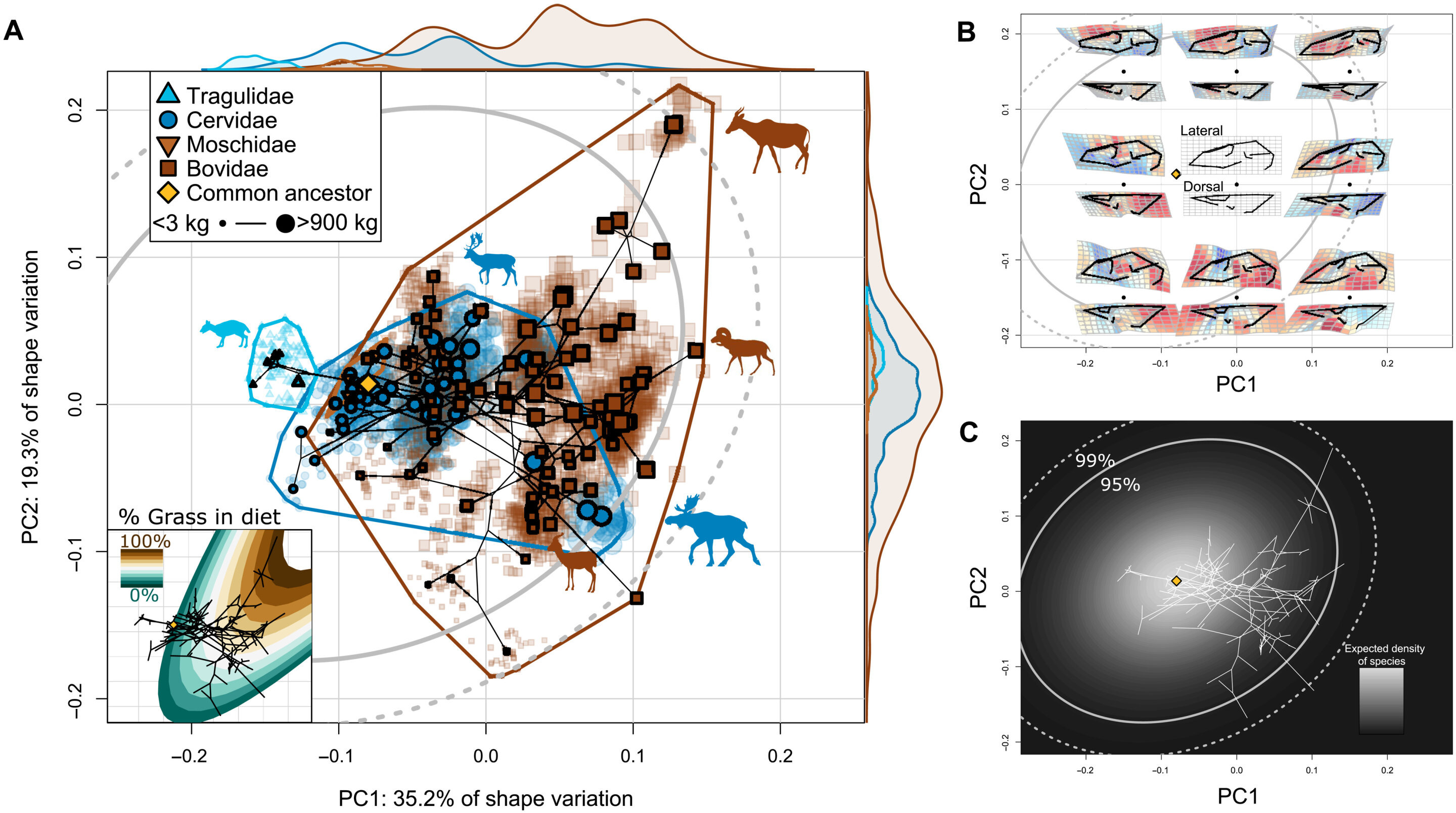
反芻動物頭蓋骨的進化變異偏向CREA。(A)物種平均顱骨形狀的種係空間,單個標本投射回該空間。灰色橢圓表示給定進化的布朗運動模型的形態空間中物種概率分布的95%和99%置信區間。數據點(種類)的大小對應於質心大小。圖邊緣顯示的分布代表每個家族的相對密度。插圖:一個生態表麵,突出了飲食中草的百分比變化,其中放牧物種主要消耗草,而食草物種消耗較少的草。注意飲食中草的百分比變化類似於物種大小的變化。(B)形態空間中的形狀變化。每個黑點代表給定坐標的形狀模型,其側麵和背麵視圖分別顯示在上方和下方。(C)給定我們的進化速率矩陣,物種在形態空間中的概率分布。根據布朗運動進化模型,較亮的區域表示一個物種在形態空間中更有可能存在的區域。該morphospace(及其他)的交互式版本可用於非移動設備(https://Daniel rhoda . shinya PPS . io/反芻動物_Dashboard/)。Credit: Science Advances, doi: 10.1126/sciadv.ade8929
(神秘的地球uux.cn)據美國物理學家組織網(by Thamarasee Jeewandara , Phys.org):進化生物學家的目標是基於種群水平變異的比較數據集,形成微觀進化過程和宏觀進化模式之間的基本聯係。在一份關於科學進展的新報告中,丹尼爾·r·羅達(Daniel R. Rhoda)和美國芝加哥大學(University of Chicago)和傑克森實驗室(Jackson Laboratory)的進化生物學科學家團隊分析了之前發表的反芻動物(哺乳動物)顱骨數據集。
通過高度保守的顱麵進化異速生長(CREA ),結果是有偏差的,其中較大的物種表現出成比例的較長的臉。研究結果強調了這一特征是一條抗性最小的進化路線,促進了與瀏覽器-草食動物譜一致的形態多樣化。結果顯示了在群體水平上的限製如何在宏觀進化尺度上產生高度定向的表型進化模式。這項工作揭示了跨哺乳動物進化枝顱麵進化異速生長的作用。
顱麵進化異速生長(CEA)
自然選擇影響群體中的表型變異,群體的發展對選擇作出反應。最大變異量的方向被稱為最小抵抗線(LLR ),代表最大進化變化的潛在方向。如果選擇與最小抵抗線一致,生物學家預計種群會朝著適應高峰的方向直接進化。但是,如果選擇的方向是別處,那麽對選擇的響應將會朝著阻力最小的線重新排列。因此,在種群水平上,適應性景觀和物種內變異限度之間的相互作用決定了表型進化的路徑。
進化生物學家試圖解釋生物多樣性的全球模式,作為生物學研究的一個基本目標,以揭示宏觀進化模式下的微觀進化機製。他們預計這些模式會影響種群水平對宏觀進化的限製。在這項工作中,Rhoda等人提出了顱麵進化異速生長(即生物的特征隨大小而變化)對微觀和宏觀進化的影響的有力證據,這可以沿著最小阻力線進行探索,以促進形態多樣性。
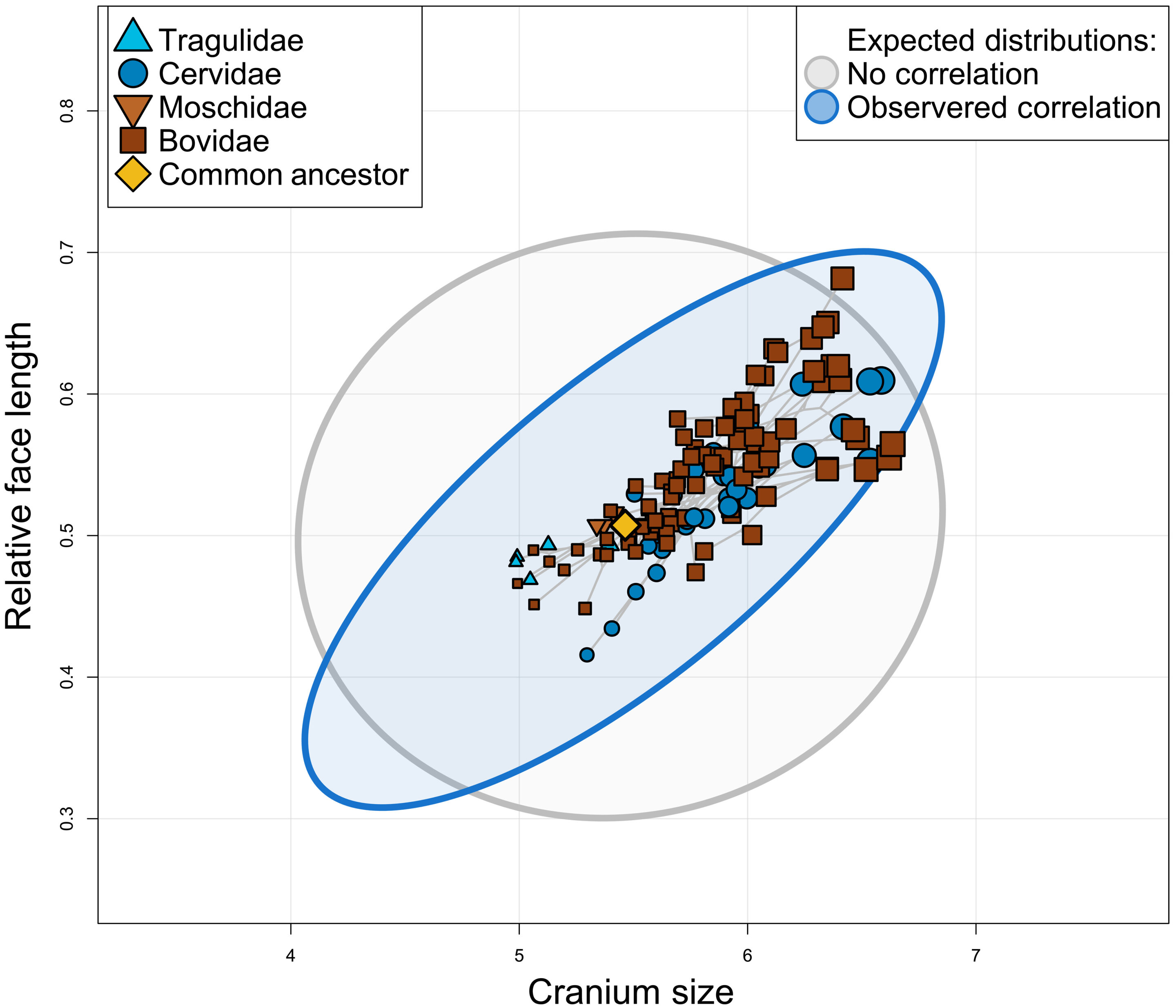
正如CREA(顱麵進化異速生長)所預測的那樣,體型較大的反芻動物有相對較長的臉。灰色橢圓代表物種預期分布的95%置信區間,如果沒有麵長的異速生長(給定觀察到的進化速率),藍色橢圓代表物種預期分布,給定觀察到的進化速率矩陣。請注意,某些“極端”特征組合(例如,非常大的長臉物種)隻能在相關進化下獲得,而其他組合(例如,非常小的長臉物種)隻能在假設的不相關進化下獲得。Credit: Science Advances, doi: 10.1126/sciadv.ade8929
哺乳動物頭骨的高度保守模式
哺乳動物頭骨保持著高度保守的個體發育和進化異速生長模式,其中較大的物種具有成比例的較長麵部,作為顱麵進化異速生長的一個例子。雖然異速生長是表型進化的一個限製,但它為極端表型的出現提供了一個沒有發育新穎性的機會。
例如,較大的短臉哺乳動物不太可能出現,而長臉、難以觸及的表型則是由於這種異速生長而出現的。因此,顱麵進化異速生長似乎是阻力最小的進化路線,簡單地表明,由於頭骨形成的尺寸限製,較大的哺乳動物有較長的臉。古生物學家注意到物種之間的大小和頭骨形狀之間有很大的關係,其中較大的物種代表較長的臉,進化異速生長的斜率在各亞科之間有很大差異。

生命穿越時間,一個視覺時間線-藝術表現的新第三紀時期-新第三紀現場。一個在蘋果樹後麵的毛茸茸的猛獁象的藝術重建,背景是早期的馬,一隻恐怖的鳥,一隻飛蟲,一隻棕色的大蝙蝠,一隻池塘裏的鱷魚,一隻岸上的劍齒貓,和森林裏的地懶。Credit: Natural History Museum, Arcata California https://natmus.humboldt.edu/exhibits/life-through-time/visual-timeline/neogene-period
物種間的頭蓋骨形狀
示例工作顯示,由於鼻內縮較大,dik-diks的顱骨形狀與其他小型反芻動物不同。然而,所有小型反芻動物的臉都比它們體型更大、臉更長的親戚要短。使用係統發育主成分分析,研究小組顯示了主導主要變異軸的清晰的大小趨勢,以進一步加強結果。Rhoda等人重建了分支之間的種間形態空間,以更好地理解不同的進化模式,同時探索顱麵進化異速生長。
該小組注意到並證實了分析中的形態空間相似性是由於異速生長——對牛科動物頭骨多樣化的主要影響。研究人員注意到牛科動物頭蓋骨進化變異的明顯大小趨勢,其中大型食草部落Alcephini和小型食草部落Neotragini相對於他們觀察到的進化速度而言,多樣化程度超過了預期。這項工作與牛科動物和鹿科動物多樣化的宏觀進化模式相一致,這種模式似乎與它們的祖先相距更遠。這些數據代表了新第三紀反芻動物飲食的進化不穩定性,同時也說明了整個新生代腐食動物多樣性的下降。
CREA在物種間的適應意義和微觀進化的動力學
Rhoda等人研究了與browser-grazer連續體相關的形狀變化,但不清楚大型放牧物種(如Alcelaphini)的異常長臉頭蓋骨是由於(1)更適合放牧的頭蓋骨的直接選擇,碰巧是長臉,還是(2)覓食生態的選擇不可知。在後一種情況下,他們的假設是,較長的臉是在與放牧相關的身體尺寸增加過程中被動進化的,這似乎不太可能,因為食物攝入和加工之間有密切的聯係。
在這兩種可能的情況下,覓食生態學為體型的進化變化提供了適應性價值,並影響了反芻動物顱骨形態的多樣化。Rhoda等人檢驗了顱麵進化異速生長(CREA)是否是反芻動物偶蹄目動物中抵抗力最低的進化係,並分析了其對形態多樣性的影響。他們直接測量與假設CREA相關的變異軸,以詢問其與種群水平進化變異的關係,並進行係統發育廣義最小二乘回歸,以測量形態差異和相對於CREA的差異角度之間的關聯。
觀點
通過這種方式,Daniel R. Rhoda和他的同事研究了哺乳動物(反芻動物)頭骨多樣化過程中宏觀進化模式背後的微觀進化過程。他們探索了顱麵進化異速生長(CREA)作為最小抵抗線(LLR)的一個方麵,因此當他們在工作中生成物種矩陣時,結果與CREA密切相關;表明這個物種比那些不太一致的物種更遠離他們的祖先。
這項工作檢查了browser-grazer譜中進化枝之間的差異及其相對於牛科和鹿科的多樣化模式的內在和外在因素。強調了內在因素對反芻動物頭骨形態多樣性的影響。這些結果廣泛地證明了內在限製對宏觀進化模式的影響,使科學家們能夠設想對CREA在其他哺乳動物分支中的作用進行進一步的研究。 |
 喜欢
喜欢 讨厌
讨厌